
目次
・カビの増殖
・冬と夏のカビの増え方の違い
・カビの増殖モデルの2Dシミュレーション

ナユミ
ひゃあ、カラーボックスにカビが生えてる!

カヤ
どれどれ、うわっ、これはすごいな…。

ナユミ
最近暑いし雨も多かったからかなあ…。壁際に置いておいたのが失敗だったかなあ。

カヤ
カラーボックスはつい壁際に置いちゃうけどな。この暑さじゃあカビも増えやすいし、仕方がないな。
ナユミ
暑いとカビ増えやすいよね…。冬は全然生えないのに、どうしてこんなに違うのかしら?

カヤ
じゃあ、試しに計算してみるかな。
カビの増殖
ナユミ
カビの増殖って、どういう計算をして表すの?
カヤ
基本的には、マルサスモデルと同じで、カビの増殖速度がカビの菌数に比例すると仮定するな。

ナユミ
人もカビも同じ数式で表すのね、なんとも言えない気分だわ。
カヤ
カビも人も生き物だから、似てるところはあるさ。ただ、カビの場合は増殖速度の温度依存性がかなりあるから、これをモデルに組み込んでみよう。
\[ \frac{dN}{dt} = k(T) N = A e^{- \frac{E}{RT}} N \]
\( N \) :菌数 \( k \) :反応速度定数 [/hr] \( A \) :頻度因子 [/hr]
\( E \) :活性化エネルギー [kJ/mol] \( R \) :気体定数 [kJ/(mol・K)]
\( T \) :絶対温度 [K]
上の式で
\[ k(T) = A e^{- \frac{E}{RT}} \]
となっていますが、この関係式をアレニウス式と呼びます。
アレニウス式を提案したスヴァンテ・アウグスト・アレニウス(1859-1927)はスウェーデンの科学者で、物理化学の創始者の一人と呼ばれているほどの大科学者です。
スヴァンテ・アウグスト・アレニウス
アレニウス式に現れる頻度因子 \( A \) は実験から定めるパラメータで、反応速度定数 \( k \) と同じ単位を持っています。
ここでは1時間あたりを採用して単位は [/hr] としていますが、1分あたり [/min] や1秒あたり [/sec] を採用しても構いません。
活性化エネルギー \( E \) も実験によって定めるパラメータで、単位にあるkJはキロジュールと読み、これはエネルギーの単位になります。
1ジュールは1ワット [W] の電力が1秒間流れたときの電気エネルギーと定められています。
なお、ジュールは日常的にもよく使うカロリーと次の関係があります。
\[ \rm 1kcal = 4.184kJ\]
また、活性化エネルギーの単位にあるmolは物質量の単位です。
物質量というのは原子や分子の個数のことですが、これらは日常の感覚からすると非常に大きな数になるため、molという塊でそれを考えます。
1mol は原子 \( 6.02 \times 10^{23} \) 個分を表します。
気体定数 \( R \) は実験とは無関係にあらかじめ定まっている値で、その値は \( 8.31451 \times 10^{-3} \ \rm [kJ/(mol \cdot K) ]\) です。
単位にある \( \rm K \) はケルビンと言って、絶対温度の単位のことです。
日本人が普通に温度と言えば、摂氏(セルシウス度 ℃)のことを指しますが、絶対温度 \( T \) [K]はこれとは目盛りがずれていて、0 [℃] は 273.15 [K] に相当します。
molと \( \rm K \) の間にある・はかけ算の記号×と同じ意味です。
モデルの微分方程式に戻ると、この式はマルサスモデルと同じ式のため解は次のようになります。
\[ N = N_0 e^{A e^{- \frac{E}{RT}}t}\]
カヤ
じゃあ、この式を使って、冬と夏でカビの増え方がどう違うのか考えてみよう。
冬と夏のカビの増え方の違い
ナユミ
冬は寒くて夏は暑いから、温度の違いを考えるのよね。
カヤ
そうだな。ナユ姉がさっき書いた微分方程式の解を二つの温度 \( T_1 \) と \( T_2 \) に適用して比をとると次のようになる。
\[ \begin{align}
\frac{N_0 e^{A e^{- \frac{E}{RT_2}}t}}{N_0 e^{A e^{- \frac{E}{RT_1}}t}} &= e^{A e^{- \frac{E}{RT_2}}t - A e^{- \frac{E}{RT_1}}t} \\\\
&= e^{\left( e^{- \frac{E}{RT_2}} - e^{- \frac{E}{RT_1}} \right) At}
\end{align}\]
菌数の比には \( N_0 \) 以外のすべてのパラメータが関わってきます。
式の形を見ると、\( A \) または \( t \) を変数とみた場合、菌数の比は指数関数となることがわかります。
\( E \) が菌数の比に与える影響を考えるために、
とりあえず、\( T_1 = 293.15 \) 、\( T_2 = 303.15 \) 、\( A = 100 \) 、\( t = 6 \) として、\( E \) を横軸にとって菌数の比をグラフにしてみました。
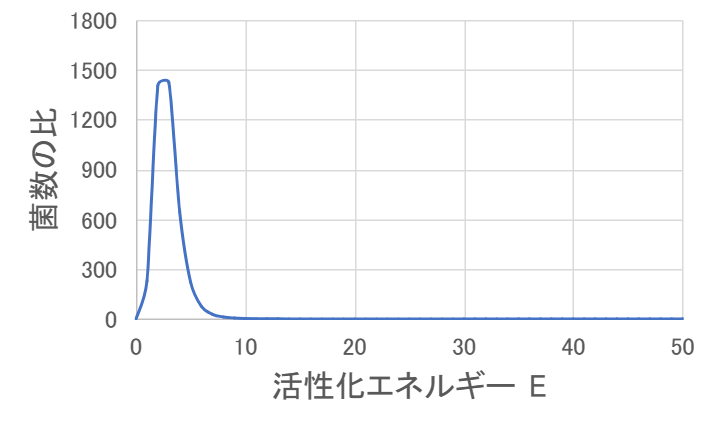
グラフにはピークがあり、それ以降は \( E \) が大きくなっても菌数の比は大きくならないようです。
そこで、このピークの位置を調べるために、次の関数を考えてみます。
\[ f(E) = e^{- \frac{E}{RT_2}} - e^{- \frac{E}{RT_1}}\]
この関数は、菌数の比の \( e \) の肩についてるかっこの中を取り出したものです。
菌数の比の \( e \) の指数部分にある頻度因子 \( A \) と時刻 \( t \) はいずれもここでは定数なので、
菌数の比のグラフが増加から減少に転じている理由はかっこの中の部分が増加から減少に転じたからとわかります。
そのため、かっこの中の関数の極大値を求めれば、そのときの \( E \) の値が菌数の比のピーク位置になっています。
そこで、極大値を求めるために、\( f'(E) = 0 \) となる \( E \) を求めてみます。
\[ \begin{align}
f'(E) &= e^{- \frac{E}{RT_2}} \cdot \left\{ -\frac{E}{RT_2} \right\} ' - e^{- \frac{E}{RT_1}} \cdot \left\{- \frac{E}{RT_1} \right\} ' \\\\
&= -\frac{1}{RT_2} \cdot e^{- \frac{E}{RT_2}} + \frac{1}{RT_1} \cdot e^{- \frac{E}{RT_1}}
\end{align}\]
1行目の式変形には合成関数の微分公式を使いました。
次は、この関数をイコール 0 とおいて、極大値を与える \( E \) を求めます。
\[ \begin{align}
0 &= -\frac{1}{RT_2} \cdot e^{- \frac{E}{RT_2}} + \frac{1}{RT_1} \cdot e^{- \frac{E}{RT_1}} \\\\
\frac{1}{T_2} \cdot e^{- \frac{E}{RT_2}} &= \frac{1}{T_1} \cdot e^{- \frac{E}{RT_1}} \\\\
\ln \left\{ \frac{1}{T_2} \cdot e^{- \frac{E}{RT_2}} \right\} &= \ln \left\{ \frac{1}{T_1} \cdot e^{- \frac{E}{RT_1}} \right\} \\\\
- \ln T_2 - \frac{E}{RT_2} &= - \ln T_1 - \frac{E}{RT_1} \\\\
\frac{E}{R} \left( \frac{1}{T_1} - \frac{1}{T_2} \right) &= \ln \frac{T_2}{T_1} \\\\
\frac{E}{R} \left( \frac{T_2 - T_1}{T_1T_2} \right) &= \ln \frac{T_2}{T_1} \\\\
E &= \frac{RT_1T_2}{T_2 - T_1} \cdot \ln \frac{T_2}{T_1}
\end{align}\]
\( f'(E) = 0 \) となる \( E \) の解がただ一つに定まったので、この式でピークの位置が求まります。
実際に先ほどのパラメータを代入してピークの位置を計算してみます。
\[ \begin{align}
E &= \frac{RT_1T_2}{T_2 - T_1} \cdot \ln \frac{T_2}{T_1} \\\\
&= \frac{8.31451 \times 10^{-3} \cdot 293.15 \cdot 303.15}{303.15 - 293.15} \cdot \ln \frac{303.15}{293.15} \\\\
&= 2.478506 \ldots
\end{align} \]
ナユミ
グラフと比較してみると…、合ってそうね。ちゃんと求まるものなのね。
カヤ
数理モデルの性質を調べる方法として、まずは計算してグラフにしてみるというのが基本だが、こういう風に式をうまく変形して特徴的な性質、今回であればピークの位置だが、そういったものを調べるというのも一つの方法だな。
ナユミ
色々な角度から眺めるのが大切なのね。
カヤ
そういうことだな。\( T_1 \) と \( T_2 \) についてはやらないが、興味のある人は同じようにピークの位置など調べてみてくれ。
それでは、菌数の比が冬と夏でどうなるかを考えてみます。
冬は10℃、夏は30℃と想定して、\( T_1 = 283.15 \) [K]、\( T_2 = 303.15 \) [K] とします。
活性化エネルギー \( E \) については参考[1]に記されている清酒酵母の活性化エネルギーの値 \( E = 54.392 \) [kJ/mol] を使います。
カビのデータではありませんが、酵母とカビは同じ真菌類に属するので、とりあえずこの値を採用します。
頻度因子 \( A \) についても、参考[1]にある清酒酵母の頻度因子の値 \( A = 1.13 \times 10^9 \) [/hr] を使います。
これらのパラメータの値を使って、時刻 \( t \) を横軸に菌数の比をグラフにしてみると次のようになります。
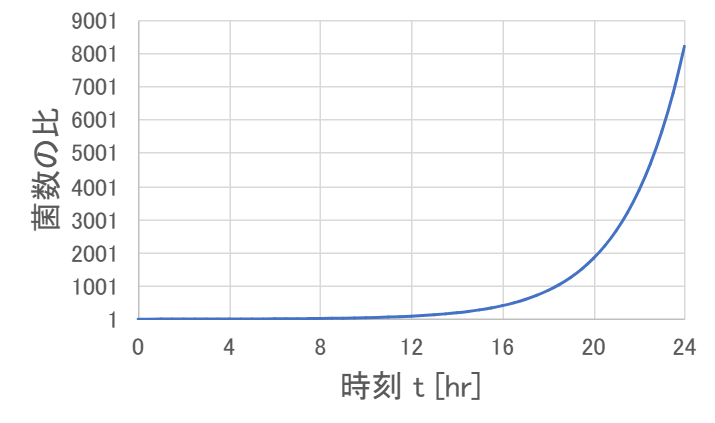
30℃で増殖したときの菌数は、24時間後、つまり1日後には10℃で増殖したときの菌数の約8000倍になっています。
ナユミ
うん、これくらいの差がつくなら、冬はカラーボックスにカビが全然生えないけど、夏はあっという間に生えるのも納得かも。
カヤ
それじゃあ最後に、少し変わった方法を使って、このモデルを二次元でシミュレーションしてみよう。
カビの増殖モデルの2Dシミュレーション
ナユミ
モデルの式って
\[ N = N_0 e^{A e^{- \frac{E}{RT}}t}\]
のこと?これでどうやって2Dシミュレーション?するの?
カヤ
色々やり方はあるんだと思うが、今回はカビの増え方を次のように仮定してみよう。
仮定1:カビは二分裂によって増える。つまり、一つの菌が二つの菌に分かれるという増え方で増える。一つの菌が三つや四つに分かれることはない。
仮定2:生存している菌はすべて同じタイミング、同じ周期で分裂する。
仮定3:菌は死滅しない。
仮定4:菌は同じ場所にいくらでも生存することができる。場所の取り合いは起こらない。
仮定2、仮定3、仮定4はあまり現実的ではありませんが、単純化のため今回はこれらの仮定を用います。
これらの仮定の下で、菌が分裂する周期を \( L \) とおき、始めの菌数を \( N_0 \) とすると、時刻 \( t \) における菌数 \( N \) は次の式で表されます。
\[ N = N_0 \cdot 2^{\frac{t}{L}} \]
\( L \) 時間ごとに菌数が2倍になるため、このような式になります。
この式は先の微分方程式の解とは一見違うようですが、
実は周期 \( L \) を適切に定めると同一の式になります。
二つの式が等しいとおいて、式を変形してみます。
\[ \begin{align}
N_0 e^{A e^{- \frac{E}{RT}}t} &= N_0 \cdot 2^{\frac{t}{L}} \\\\
e^{A e^{- \frac{E}{RT}}t} &= 2^{\frac{t}{L}} \\\\
A e^{- \frac{E}{RT}}t &= \ln 2^{\frac{t}{L}} \\\\
A e^{- \frac{E}{RT}}t &= \frac{t}{L} \cdot \ln 2 \\\\
A e^{- \frac{E}{RT}} &= \frac{1}{L} \cdot \ln 2 \\\\
L &= \frac{\ln 2}{A} \cdot e^{\frac{E}{RT}} = \frac{\ln 2}{k(T)}
\end{align}\]
最後の式の形は半減期の式と同じです。
ただ、今回は崩壊反応ではなく増殖反応なので、ここでの周期 \( L \) は倍加時間と呼ばれています。
ナユミ
この式を使って2Dシミュレーションをするのかな?
カヤ
そうなんだが、もう少し準備が必要だな。菌が分裂する空間とその分裂の仕方について、次のように定めておこう。
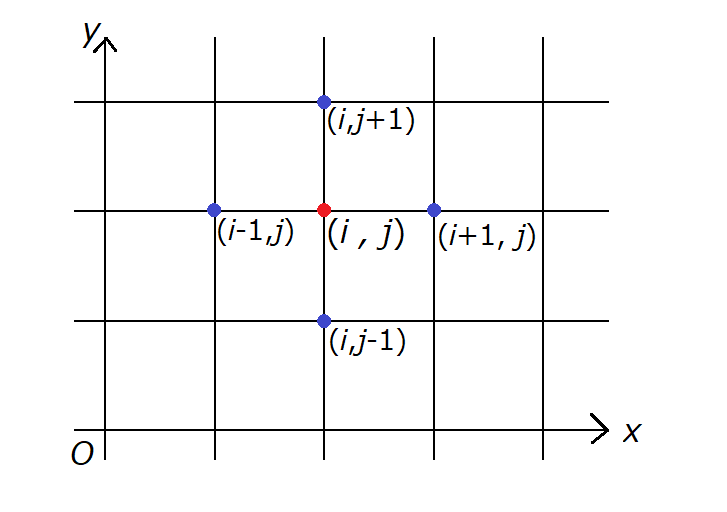
菌が分裂する空間は \( xy \) 座標平面上の \( x \) 座標および \( y \) 座標がいずれも0以上の整数となっている点 \( (i,j) \) に限定する。
菌は時刻 \( t = 0 \) に分裂した後、\( L \) 時間ごとに1回分裂する。分裂した菌の増加分は四方のいずれかに等しい確率で移動する。
四方のいずれかに等しい確率で移動するというのは、点 \( (i,j) \) に居る菌が分裂して増えた分は、点 \( (i+1,j) \) 、点 \( (i,j+1) \) 、点 \( (i-1,j) \) 、点 \( (i,j-1) \) のいずれかに移動し、それぞれの場所への移動確率は \( \frac{1}{4} \) であるという意味です。
下図で表すと、赤丸で分裂した菌のうち増えた分が4つの青丸のどれかに移動するということです。
カヤ
ここまでの設定で2Dシミュレーションを行うことができる。下にシミュレーションマシンを作ったから遊んでみてくれ。パラメータの値を変更したいときは、ブラウザでこのページを再読み込みしてからやり直してくれ。
0hr
| |
|
|
|
|
|
| 1未満 |
1 |
10 |
100 |
1000 |
10000以上 |
ナユミ
また面白いものを作ったわね…。10℃と30℃でカビの増え方に差があるのがよくわかるわ。でも、カビというより苔玉みたいね。
カヤ
まあ、グラフィックの拙さは許してくれ。カビの繊維っぽさが表現できるとよかったんだがな。

ナユミ
でも、これくらいのほうが手作り感があっていいかもしれないわよ。

カヤ
だと嬉しいな。
参考:
[1] 永谷正治、<連載講座> 第11章 微生物の反応速度、醸造工学、第68巻 第11号、1973年
[2] Wikipedia スヴァンテ・アレニウス、https://ja.wikipedia.org/wiki/スヴァンテ・アレニウス、2023年8月17日閲覧
[3] 池上雄作・岩泉正基・手老省三、化学教科書シリーズ 第2版 物理化学Ⅱ―熱力学・速度論―、丸善、1996年12月20日発行
[4] 海野肇・中西一弘 監修、丹治保典・今井正直・養王田正文・荻野博康 著、生物化学工学 第3版、講談社、2011年9月20日発行
[5] Wikipedia 倍加時間、https://ja.wikipedia.org/wiki/倍加時間、2023年8月20日閲覧
前の記事
第3話
複利計算
次の記事
第5話
ロジスティック曲線